The chemical research on the plant cannabinoids and their derivatives over two centuries is concisely reviewed. The tortuous path leading to the discovery of the endogenous cannabinoids is described. Future directions, which will probably be followed are delineated.
Raphael Mechoulam *, Lumı´r Hanus
Department of Medicinal Chemistry and Natural Products, Hebrew Uni6ersity of Jerusalem, Medical Faculty, Ein Kerem Campus, 91120 Jerusalem, Israel Received 27 April 2000; received in revised form 5 June 2000; accepted 5 June 2000
1. Introduction
About 150 years ago the Pharmaceutical Society of Paris awarded a prize to Personne (1855) for ‘a good analysis of hemp’ (Robiquet, 1855). Unfortunately Personne’s volatile oil was shown later to be an inactive, impure sesquiterpene (Vignolo, 1895). Actually research on the chemistry of Cannabis had already begun decades earlier, as a major trend in chemical research throughout the nineteenth century was the quest for active natural products. Numerous alkaloids were isolated in pure form from various plants and some of them were partially characterized. Morphine, cocaine, strychnine, and many others were purified and used in medicine. However, most of the major terpenoids were not isolated until the end of the century or even much later, and in many cases their purity was doubtful. The reason behind this disparity is that alkaloids are relatively easy to separate and crystallized as salts, whereas terpenoids are usually present in mixtures whose separation is tedious and in many cases was impossible with the techniques available to the chemist of a 100 years ago. The search for the active component of Cannabis sati6a is probably one of the best examples. Numerous groups reported initial efforts to obtain an active Cannabis component after news of the extensive medical useof its resin in India and the East were widely circulated and the Parisian literary circle around Baudelaire indulged in hedonistic use of Middle Eastern hashish. However, more than a century * Corresponding author. Tel.: 972-2-6758634; fax: 972-2-6410740. E-mail address: mechou@yam-suff.cc.huji.ac.il (R. Mechoulam).
passed until the major active component, D9 - THC, was isolated in pure form and its structure was elucidated. For a detailed history of early Cannabis research see Mechoulam (1973). Schlesinger (1840) was apparently the first investigator to obtain an active extract from the leaves and flowers of hemp. A few years later Decourtive (1848) described the preparation of an ethanol extract that on evaporation of the solvent gave a dark resin, which he named ‘cannabin’. A resin prepared in a similar manner (called ‘hashishin’) was used by Gastinel (1848) as a drug. Work of closely similar nature was reported by Robertson (1847), Savory (1843), while Smith (1885), Smith and Smith (1847) ascribed the physiological action to a resin that they obtained by alcoholic extraction of the dry plant. To the solution a milk of lime as thick as cream’ was added to remove chlorophyll. After filtration and treatment with sulfuric acid the solvent was evaporated. The resin was not basic (i.e. an alkaloid) but neutral. This was unexpected since most chemists at the time assumed that the active principle was, such as those of opium and cinchona, an alkaloid. The authors record that ‘two-thirds of a grain of this resin acts upon ourselves as a powerful narcotic, and one grain produces complete intoxication. In this character it is quite analogous to alcohol, but in its hypnotic and soothing effects on the nervous system its resemblance to morphia is very great’.
1.1. Alkaloids
For many years during the 19th century the possible presence of alkaloids was discussed. Preobrajensky (1876) claimed the presence of nicotine in Cannabis sati6a resin but this was refuted by Kennedy (1886). Hay (1883) was able to obtain an alkaloid, tetanocannabin, which possessed strychninelike properties when tested on a frog. Although the balance of evidence was infavor of some alkaloidal substance (in addition to choline and trigonellin) being present in small quantity in Indian hemp, almost no further work along these lines was undertaken. This lack of data apparently did not prevent the marketing of ‘Cannabine Alkaloid Merck’ (Merck Index, 1896), as ‘fine needles … hypnotic without dangerous secondary effects’. The first solid indication of the presence of alkaloids in Cannabis , in addition to the abovedescribed simple nitrogenous bases, was put forward much later in Klein et al. (1971). Alkaloidal mixtures were extracted from plant material. The total crude yield was 0.02%. Four alkaloids, named cannabimines A–D, were isolated by preparative TLC. High-resolution mass spectrometric data indicated molecular weights of 279– 363. Structural assignments could not be made. No further work on these alkaloids has been reported. Cannabimine A, C21 H37 N3 O2 may be identical to anhydrocannabisativine, which shares the same molecular formula. The first Cannabis alkaloid to be fully identified was cannabisativine which was isolated in Mississippi in 1975 from the roots of a Mexican variant (Lotter et al., 1975). TLC indicated the presence of this substance in the leaves of the plant as well. The structure of cannabisativine was elucidated by X-ray crystallography. A second alkaloid, named anhydrocannabisativine, was isolated in a miniscule yield by the same group from the leaves and roots of a Mexican variant of Cannabis sati6a (ElSohly and Turner, 1976). Anhydrocannabistivine has been found in plant samples of Cannabis from 15 different geographical locations. Several related spermidine alkaloids palustrine and palustridine (both found in Equisetum species) are known in nature. While the crude alkaloidal mixture caused decreased activity in mice, in view of the low concentration of the alkaloids in the plant, it is doubtful whether these components are relevant to Cannabis activity in man. For a detailed review of cannabinoid alkaloids see Mechoulam (1988).
2. Cannabinoids
At the turn of the 19th century a group in Cambridge (Wood et al., 1896) effected a considerable purification of the components of Cannabis by fractional distillation of an ethereal extract from charas. They obtained a terpene and a highboiling, viscous oil. The viscous oil (‘amber colored when seen in thin layers but ruby red when seen in mass’) was assumed to be a single substance. The fractions isolated by the Cambridge group were tested by Marshall (1897, 1898). In the Wood et al. (1896) article he is quoted as follows.
The red oil, is extremely active, and taken in doses of 0.05 g induces decided intoxication followed by sleep. The symptoms produced by it are peculiar to Cannabis indica , and as none of the other products appear to possess this action, this substance must be regarded as the active constituent of the plant. Later Dunstan and Henry (1898), Wood (1899) found that the oil was not homogenous and isolated from it, after acetylation, a crystalline acetate which could be hydrolyzed to a resinous phenol that analyzed for C21 H28 O2 . To this component they gave the name cannabinol. Its lack of optical rotation (in contrast to the negative values of the other major natural cannabinoids known today) and the crystalline cannabinol acetate obtained from cannabinol emphasize its purity. Thus, cannabinol represents the first natural cannabinoid to be obtained in pure form. Pure cannabinol apparently was not tested at that time for its biological activity. Hence, on the basis of work on the crude extract (Marshall, 1898; Fraenkel, 1903) it was wrongly assumed that this component was the active principle of Cannabis .
The crystalization of cannabinol acetate from the red oil indicates that cannabinol was present in it in relatively high concentrations. Wood may had worked with old, deteriorated samples of hashish in which much of the active constituent tetrahydrocannabinol (THC) had oxidized to cannabinol (see below). We know today that cannabinol is a relatively minor constituent in fresh Cannabis , hashish, or charas and may in fact be an artifact. All the major cannabinoids boil in the same temperature range, and their separation is impossible by distillation. Indeed for many years numerous groups unsuccessfully tried to repeat and expand Wood’s findings, hence the work of the Cambridge group was largely ignored.
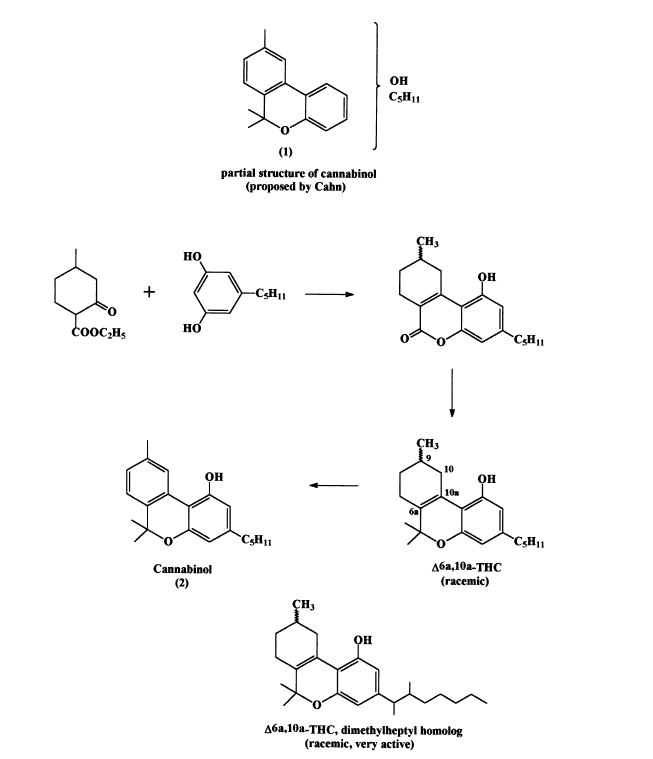
As late as 1927 it was claimed that ‘cannabinol’ has a rotation of 150° (Casparis and Bauer, 1927), although Wood has already shown that it lacks rotation. In the early 1930’s Cahn again obtained pure cannabinol, reinvestigated and expanded the chemical degradations reported earlier by Wood (Cahn, 1933 and papers cited). In the 1930’s the only available route for structural elucidation of a novel natural product was by chemical degradation to known compounds, followed by a crossword- type effort to establish the structure. On the basis of the extensive chemical degradations Cahn arrived at the conclusion that cannabinol has the partial structure 1 (see Scheme 1). Pure cannabinol was found to be weakly active in a dog ataxia test and the quest for the active component(s) continued in the late 1930’s and early 1940’s, mainly in the laboratories of Roger Adams in the US and A. Todd (later Lord Todd) in UK. Apparently the early efforts were not successful and both groups then followed two parallel paths in their research, syntheses of the various cannabinol isomers, suggested by Cahn’s partial structure, and further efforts to obtain the natural active constituent(s). Several of the possible structural isomers of cannabinol were indeed prepared. One of the synthetic compounds (2) was identical to cannabinol isolated by Cahn thus establishing its structure (Adams et al., 1940; Adams, 1941–1942; Todd, 1946).
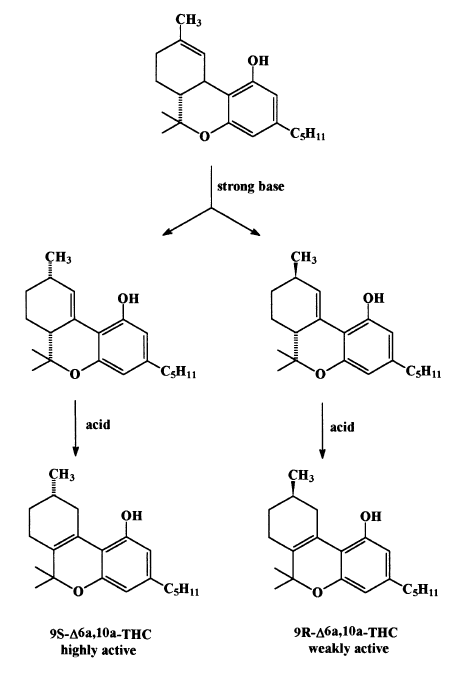
The synthetic route is presented in Scheme 1. The tricyclic intermediates in the cannabinol synthesis were tested for cannabinoid activity in the dog ataxia assay and the racemic D6a,10a -tetrahydrocannabinol was found, quite unexpectedly, to be active. As the natural D9 -THC has not been isolated at that time, this racemate was used in numerous pharmacological investigations, particularly by Loewe (1950), who collaborated with R. Adams. About 40 years later the two pure enantiomers of the racemate, namely the C9 -R(4) and C9 -S(5), were synthesized (Scheme 2, Srebnik et al., 1984) and assayed in humans for psychoactivity (Hollister et al., 1987). The C9 S enantiomer had definite psychotropic actions, qualitatively similar to those of D9 -tetrahydrocannabinol, but quantitatively much less potent. Adding the two enantiomers together did not increase the effect, confirming that activity was solely in the one enantiomer and that there was no interaction between them.
For nearly a decade (1940–1949) the Adams group published the preparation and evaluation of derivatives of D6a,10a -THC. Of particular importance and interest was the discovery that the THC homolog in which the pentyl side chain was replaced by a 1,1-dimethylheptyl or a 1, 2-dimethylheptyl one was several hundred times more active than the original pentyl compound (Adams et al., 1949). For structure, see Scheme 1.
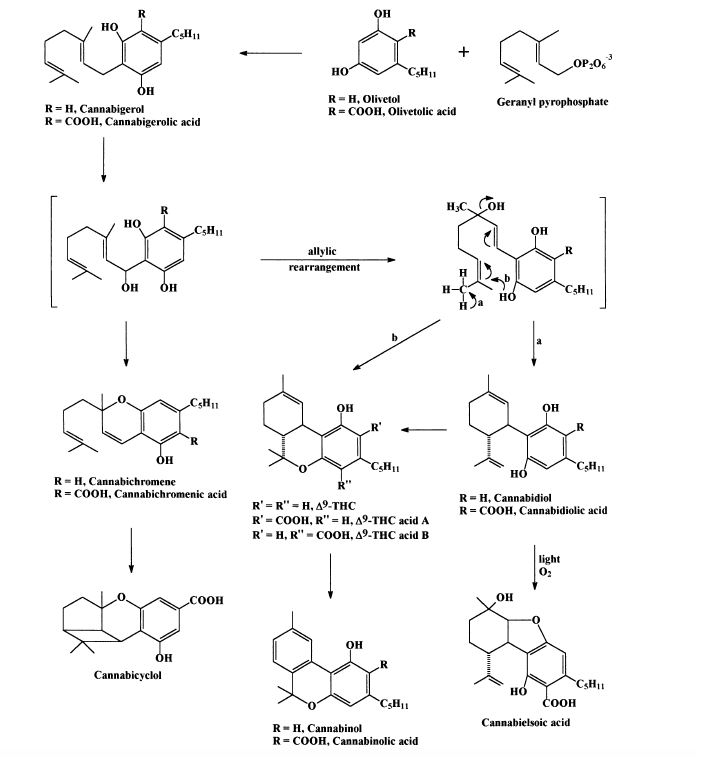
The first isolation in a pure form of a psychoactive Cannabis principle, D9 -THC, was reported finally in 1964 (Gaoni and Mechoulam, 1964, 1971). A hexane extract of hashish was separated into acidic and neutral fractions. Repeated chromatography of the neutral fraction on Florisil, acid-washed alumina, and alumina containing 12% silver nitrate eluted the following compounds (in order of increasing polarity), a mixture of waxy, noncannabinoid materials, cannabicyclol, cannabidiol, D9 -THC, cannabinol, cannabichromene, cannabigerol, and polar constituents and polymers. All cannabinoids were obtained in crystalline form, except D9 -THC and cannabichromene, of which crystalline derivatives (D9 -THC, 3,5 dinitrophenylurethane, m.p. 115–116°C; cannabichromene, 3,5- dinitrophenylurethane, m.p. 106–107°C) were prepared. On hydrolysis pure D9 -THC and cannabichromene were obtained. For structures see Scheme 3.
Cannabidiol had already been obtained in the early 1940’s, but its structure and stereochemistry were determined only in 1963 (Mechoulam and Shvo, 1963) mainly on the basis of the then novel NMR method. The elucidation of the structure of CBD was of considerable importance as it was on its basis that the structure of THC was elucidated, again mostly byNMRanalysis.Afinal proof of the structure was made by the conversion of CBD into THC by a mild acid treatment. The absolute configuration of D9 -THC was established in (Mechoulam and Gaoni, 1967). The activity of THC was initially established by evaluation in rhesus monkeys (Grunfeld and Edery, 1969; Mechoulam et al., 1970). Since then thousands of papers have been published on the activity of D9 -THC, in vitro and in vivo, including human trials. The structures of cannabigerol, cannabichromene, cannabicyclol, and a long list of additional, nonactive cannabinoids were established over the next decade. Of particular interest are the cannabinoid acids which seem to be the original cannabinoids formed in the plant but are later decarboxylated (possibly in part in the plant itself) to yield the better known neutral cannabinoids described above.
For a detailed review of the natural cannabinoids see Turner et al. (1980). Both Adams and Todd (Adams, 1941–1942; Todd, 1946) also obtained a further inactive component which was named cannabidiol. Its structure was not fully determined although it was obviously a major constituent. An isolated report on the active principle appeared in De Ropp (1960). He obtained an oily component, presumed to be a tetrahydrocannabinol, although the analytical values presented did not support this conclusion. However, the positive dog ataxia test tends to substantiate the assumption that a THC was obtained. The purity of this material, however, is difficult to determine. No structural work on this component was reported.
Leaves of Cannabis sati6a , var. indica, cultivated in Czechoslovakia were investigated by Krejcˇı´ (1952) at the Palacky´ University at Olomouc in Czechoslovakia. He found that an ethyl alcohol extract was active against gram-positive microorganisms. The S& antavy´ group also prepared an acetyl derivative of the compound responsible for these effects (Krejcˇı´ and S& antavy´, 1955) and the substance was named cannabidiolic acid on the basis of its conversion to cannabidiol. It was the first cannabinoid acid isolated from hemp. The antibacterial effect was confirmed in in vivo (topical) and in vitro assays (Kabelı´k et al., 1960).
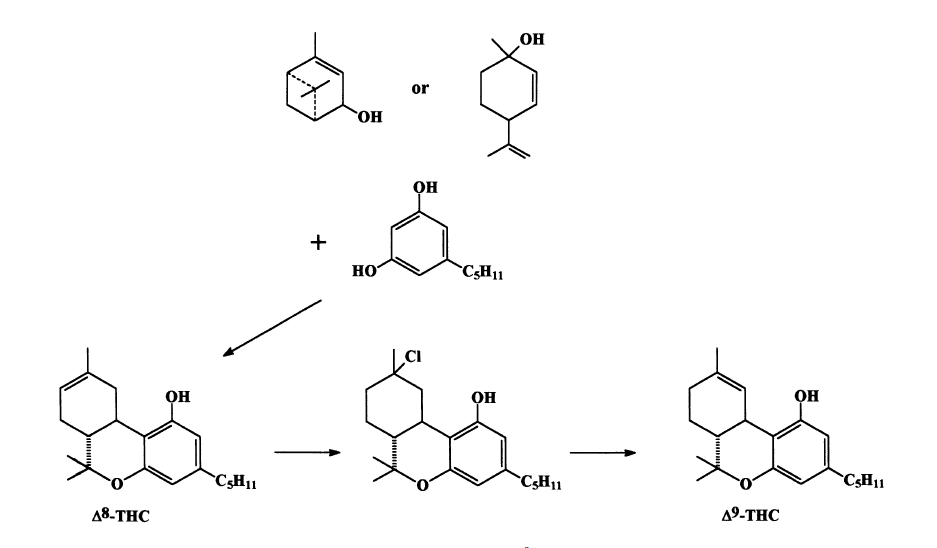
The great abundance of cannabinoids has made possible a reasonable suggestion for the biogenesis of the cannabinoid family of plant constituents (see Scheme 3). Some recent papers have given support to the proposed biogenetic pathway. Of particular interest is the unexpected discovery by Taura et al. (1995) that cannabigerolic acid does not give, after oxidation, cannabidiolic acid but may proceed directly to D9 -THC acid A. The next step undertaken by several groups was the synthesis of the natural cannabinoids. The first total synthesis was reported in 1965 and later two synthetic routes, which are still being used, were published. They are quite similar in nature: both involve the condensation of a monoterpenoid with olivetol, to yield D8 -THC (with the D9 double bond moving to the D8 position). The D8 -THC could be converted back to D9 -THC by conversion to the tertiary chloride which could be eliminated as hydrochloric acid by an internal phenolate attack (Scheme 4).
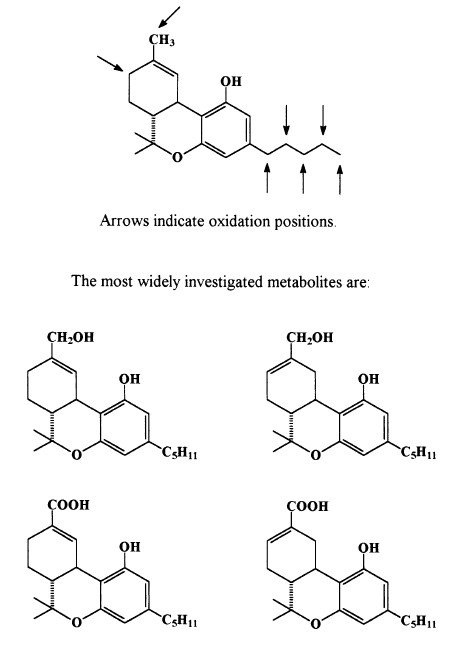
These two, by now quite ancient syntheses, are still employed for the preparation of CBD, D9 -THC (marketed as Marinol) or, via a minor modification, for HU-211 (named Dexanabinol), a drug in phase 3 clinical trials. An excellent summary of cannabinoid syntheses has been published (Razdan, 1981). The first cannabinoid metabolite to be identified was 11-hydroxy-D8 -THC (Ben-Zvi et al., 1970). As often happens in the history of science, four groups almost simultaneously identified the hydroxylation of the C-11 position as the first metabolic step (Ben-Zvi et al., 1970; Lemberger et al., 1970; Nilsson et al., 1970; Wall et al., 1970, Scheme 5). Both 11-hydroxy-D8 -THC and 11-hydroxy- D9 -THC were reported within a period of several months. 11-Hydroxy-D8 -THC was soon synthesized (Ben-Zvi et al., 1970) and was shown to be as active as D8 -THC itself.
The next metabolic step was shown to be the conversion of the allylic hydroxyl group into a carboxyl group (Scheme 5). The so formed THC- 7-oic acid was inactive (Mechoulam et al., 1973). It was shown to remain in the body as a glucoronide over many weeks. For a review of cannabinoid metabolism see Agurell et al. (1986). By the mid 1970’s the chemical research on the plant cannabinoids seemed to have reached its goals: most cannabinoids had been isolated and synthesized and the metabolic pathways had been elucidated, with the metabolites identified and synthesized. Some industrial firms and academic aboratories initiated projects to develop cannabinoid- based drugs (see Milne et al., 1981). However, separation between the psychotropic effects and the medically useful ones was not achieved and except for Nabilone (Archer et al., 1986), a potent THC-type drug, success was elusive. Nabilone is used in some countries, UK for example, as an antiemetic agent. D9 -THC itself is also used for this purpose as well as for enhancement of appetite, particularly by AIDS patients (Mechoulam et al., 1998a). D8 -THC, which is considerably less expensive to prepare, more stable and as active as D9 -THC in antiemetic studies (Abrahamov et al., 1995) is not marketed, apparently for purely commercial reasons.
3. Endocannabinoids
Contrary to the advances made in the chemistry of cannabinoids the molecular basis of cannabinoid activity remained an enigma for several decades. During the 1970’s and early 1980’s it was generally assumed that the high lipophilicity of the cannabinoids is the basis of their pharmacological action. D9-THC was considered to belong to the group of biologically active lipophiles and that its effects should be compared with the chronic effects of anaesthetics and solvents (Paton, 1975). Thus, it seemed possible to explain the action of cannabinoids without postulating the existence of specific cannabinoid receptors and of endogenous mediators. Gill and Lawrence (1976) found experimental evidence to support the above suggestion and concluded that, ‘… as the liposomal membrane is apparently able to discriminate between the various cannabinoids in a way similar to the nerve cell, it is unnecessary to postulate the existence of a more complex macromolecular receptor substance to account for the observed structure-activity relationships’.
A further, conceptual, problem which hampered work aimed at the discovery of a cannabinoid receptor system was the presumed lack of stereospecificity. As receptors, which are proteins, are asymmetric, binding to a receptor should take place only with one of the two asymmetric components of a racemate. Early work indicated that some Cannabis-type activity was observed with the unnatural ()D9-THC. This excluded the possibility of a receptor-type activity of cannabinoids. However, by the mid 1980’s work by several groups, including ours, indicated that cannabinoid action was highly stereospecific contrary to previous work which had apparently been done with samples which contained ( )D9 - THC as impurity. Thus, the very potent synthetic cannabinoid HU-210 is several thousand times more active than its synthetic mirror image (HU- 211) in many tests in animals, as well as in biochemical ones (Mechoulam et al., 1988). These results prompted several groups to look for a specific receptor in brain and on neuronal cells, which would bind the psychoactive cannabinoids. In 1988 Howlett’s group in St. Louis established the presence in rat brain of a specific cannabinoid receptor by the use of a tritium labeled cannabinoid (Devane et al., 1988). It is known today as CB1 . This receptor has been cloned (Matsuda et al., 1990).
It was quite unacceptable to most neuroscientists that the brain will waste its resources to synthesize a receptor in order to bind a constituent of a plant. The only reasonable assumption, which could be made was that the brain must produce a neuronal mediator, a specific compound (or a family of compounds) which binds to and activates the cannabinoid receptor. The plant cannabinoid, THC, by coincidence happens to bind to the same receptor. In the late 1980’s several groups initiated work aimed at the discovery of such a brain constituent. None of many known hormones, neurotransmitters or other biologically active compounds of various types were found to bind to the receptor. Obviously a new type of ligand was involved. Evans et al. (1992) examined the ability of a calcium ionophore to release compounds that can interact with the cannabinoid receptor. They found that the ionophore A-23187 released an unidentified material with receptor-binding activity in the presence of Ca ions. However, this material was not isolated and its structure is unknown.
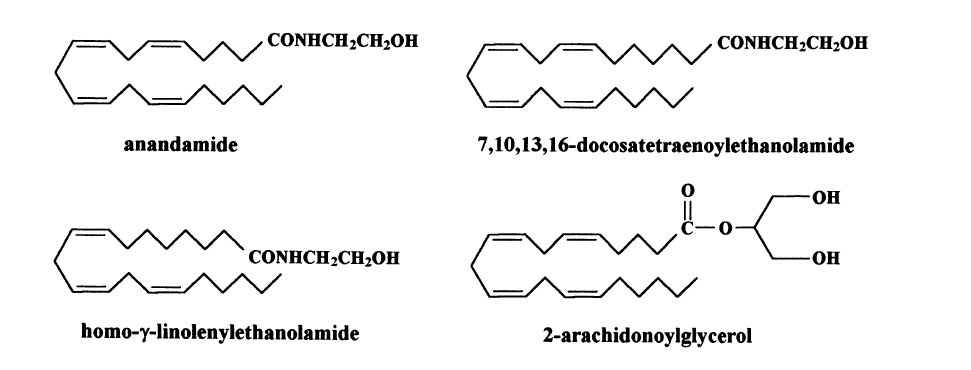
Our group proceeded by preparing a novel, highly psychoactive probe for the cannabinoid receptor, which could be easily labeled with tritium (Devane et al., 1992a). This probe binds to the receptor and the formed complex was used to assay brain fractions for Cannabis -like activity. A lipid-soluble fraction isolated from pig brain exhibited Cannabis -like activity: on incubation with the receptor, it displaced the radioactive probe. Purification of the constituents of this fraction over many different chromatographic columns ultimately led to the isolation of a single active constituent. With advanced physical techniques of NMR spectroscopy and mass spectrometry, it was possible initially to establish that the endogenous brain constituent was a fatty acid derivative, and then to elucidate its complete structure as arachidonoylethanolamide (Devane et al., 1992b, Scheme 6). The final proof of the suggested structure was its synthesis from readily available materials. We named this endogenous brain constituent ‘anandamide’ based on the Sanscrit term ‘ananda’ meaning ‘bliss’ and the amide moiety present in the novel compound. When tested in animals anandamide paralleled THC as regards analgesia, sedation, motor coordination, and certain biochemical parameters. Later, using the same techniques two additional compounds, structurally closely related to anandamide, were isolated from porcine brain (Hanusˇ et al., 1993, Scheme 6). Both bind to the cannabinoid receptor. The brain apparently produces a whole family of anandamide-type compounds.
A second cannabinoid receptor (CB2 ) has been identified in and cloned from rat spleen. CB2 has so far been found mostly in cells of the immune system (Munro et al., 1993). Using the methods used by our group for the identification of anandamide we identified a cannabinoid ligand from canine gut, which binds to both CB2 and CB1 . Its structure was elucidated as 2-arachidonoylglycerol (2-AG) (Mechoulam et al., 1995, Scheme 6). Later independently, Sugiura et al. (1995) also published the identification of 2-AG as an endocannabinoid and established its presence in the brain. Both groups had been presenting their work in lectures before the publications but, surprisingly, were unaware of the parallel research going on in the laboratories of their colleagues. 2-AG being an ester is a labile compound, easily hydrolysed in the body (for a review see Mechoulam et al., 1998b).
Apparently a rather ingenious way of reduction of the rate of its hydrolysis, and thus of enhancement of its activity, in the body is by the formation and presence alongside 2-AG of large concentrations of related, inactive, fatty acid esters of glycerol (an ‘entourage’ effect). Indeed 2-AG is always accompanied by palmitoyl glycerol and linoleyl glycerol, present in concentrations much higher than that of 2-AG (Ben-Shabat et al., 1998). The Ki of 2-AG to CB1 in the presence of the entourage compounds is much lower than when assayed alone. The in vivo effects of 2-AG are also more potent. The possible ‘entourage’ effect in the anandamide series has not yet been looked into.
The structural activity relationships in the tricyclic as well in the fatty acid series have been the object of a long list of publications, but this aspect of research is beyond the scope of the present review (Mechoulam et al., 1992; Sheskin et al., 1997; Martin et al., 1999). It is possible toobtain cannabinoid ligands that bind specifically to one of the receptors only. Thus HU-308, a synthetic compound, binds specifically to CB2 (Hanusˇ et al., 1999). Antagonists specific for CB1 and for CB2 have also been reported and are widely employed (Barth, 1998). Also, anandamide derivatives which are more stable to enzymatic attack than anandamide have been prepared (Abadji et al., 1994). The endocannabinoids, in particular anandamide, have been the object of many hundreds of publications, in chemistry, biochemistry, physiology and pharmacology. They will be reviewed in additional papers in this Symposium-in-print. However no endocannabinoids have been administered to humans so far. Although they are natural products, their toxicology is not yet known and hence according to present day regulations they cannot be evaluated in humans.
4. Future directions
In what direction will chemical research in the field of cannabinoids proceed? An ancient Jewish saying states that prophecy powers are given to fools only. With the hope that we shall not be exposed as fools, we believe that chemical research over the next decade will proceed, in part at least, along the following paths.
1. Identification of novel receptor sites and ligands.
2. The elucidation of the 3 dimensional structures of plant, synthetic and endocannabinoids for binding to the receptors, and the differential requirements for the two known receptors.
3. Novel chemical approaches to specific drugs against pain, inflammation, multiple sclerosis, brain damage, etc.
4. Chemical–physiological investigations on novel types of actions of the endocannabinoids. The ultimate goal of such investigations could be the possible involvement of the endocannabinoids in the chemistry of emotions — a field still an enigma.
Acknowledgements
Research in the laboratory of the authors was supported by NIDA.
View Larger
Scheme 1. Adams and Todd synthesis of cannabinol.
View Larger
Scheme 2. Synthesis of enantiomeric D6a,10a-THC’s.
View Larger
Scheme 3. Some natural cannabinoids and their assumed biogenesis.
View Larger
Scheme 4. Synthesis of D9-THC.
View Larger
Scheme 5. Metabolism of D9-THC.
View Larger
Scheme 6. The known endocannabinoids.
